染色体の真の姿を
紐解く挑戦

- 京都大学
大学院理学研究科
ゲノム多元統御学
教授 - 西山 朋子
学術変革領域(A)「ゲノムモダリティ」が発足した。本領域は、新学術領域「染色体オーケストレーションシステム(染色体OS)」(H27-H31)の後継として、染色体OSで得られた染色体構造と機能の連携機構に、さらに「DNAの高分子物性理解」という観点を加え、生物物理学分野への先端的な発展を図る領域である。DNA分子の物性は、分子生物学の勃興期にこそ着目されていたものの、情報科学としてのゲノム解析の爆発的な発展のなかで忘れ去られていった。しかし、ヌクレオソームの動態、クロマチンの高次な折りたたみによる遺伝子発現制御、分裂期染色体の構築等の諸機能を真に理解するためには、DNAの構造物性的側面を深く理解しなくてはならない。
従来のゲノム研究では、塩基配列を基盤としたゲノム制御と非塩基配列情報を基盤としたエピゲノム制御という二つの強力な概念を軸に、ヌクレオソーム構造を基本としたゲノムの構造と機能の実体が明らかにされてきた。これまでのゲノム研究の潮流は、まさにこの二つの概念によって方向付けられてきたと言っても過言ではない。こうした従来のゲノム研究の潮流に、近年、全く新しいゲノム構造構築の概念が加わろうとしている。その一つの例がループ押出し(loop extrusion)機構によるDNAループの形成である。Hi-Cをはじめとした新規ゲノム構造解析技術の急速な普及を背景に、間期クロマチンと分裂期染色体におけるループ様ドメイン構造の存在が明らかされ、これらの構造の形成にSMCタンパク質複合体が重要な役割を果たすことが分かってきた。これは非ヌクレオソーム型染色体構造構築の一例である。また、古典的な例を挙げれば、精子核クロマチンも非ヌクレオソーム型クロマチンだが、そのDNA折りたたみのメカニズムは全く理解されていない。これら、ヌクレオソームを基本とした構造では説明できない、新しいゲノム構造構築原理を理解するために、DNA物性理解は必要不可欠である。また、ヌクレオソームにしても、そのゲノム配列上でのポジショニングや巻き付きの度合い、リンカーの長さや硬さ、DNA・ヒストンの化学修飾や各種タンパク質因子の結合がどのようにその配列・配向と動態に関与し、転写等の分子反応の制御に関わっているかなど、DNA物性との関わりは十分に理解されてはいない。
ゲノムモダリティ Genome modality とは、「塩基配列情報、DNA物性、その他の環境諸因子によって多元的に制御されるゲノムの構造や機能の様式」を意味する造語である。ゲノムという実体を、DNA物性を含む複眼的な視点で捉えることが、本領域の挑戦である。一分子レベルのDNAやタンパク質動態が可視化・解析できるようになった現在、改めてDNA本来の物理的性質に立ち返り、5年間の研究期間の中で、染色体の真の姿を紐解いていきたい。「塩基配列」と「ヒストンコード」に「DNA物性」を加えた、包括的な「ゲノムモダリティコード」によって、ゲノムを描き出す日が来ることを期待してやまない。
Outline of research 研究の概要
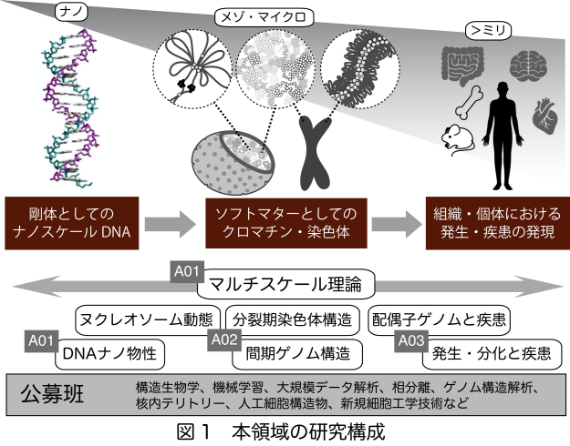
本領域では、ナノスケールのDNA・ヌクレオソームから、組織や個体にまで及ぶ幅広いスケールを研究対象とする(図1)。ゲノムモダリティを制御する要因として、DNAの物性に加えて、核内や細胞内環境、広い意味でのタンパク質物性、液−液相分離を代表とする物理化学反応等を想定し、これらの要因がそれぞれのスケールでゲノムモダリティをどのように制御し、染色体やクロマチンの振る舞いを規定するのか、その制御がどのように細胞機能に直結し、その破綻が発生異常や疾患を引き起こすのかを、理論・計測・実験的再構成・ゲノミクス等の異なる手法を用いて解明していく。本領域では、ゲノム構造の各階層に応じて三つの研究項目を設け、研究項目A01「ゲノムモダリティの理論と基盤」では、高分子物理学に基づいたナノスケールゲノム構造形成原理の追求を行うとともに、周辺環境に応じた構造・機能制御原理を理解する。またナノスケールから高次ゲノム構造に至る各階層を理論的に連結するマルチスケール理論構築を行う。研究項目A02「メゾスケールのゲノムモダリティ」では、ヌクレオソームやDNAループ構造、クロマチンファイバー/ドメイン構造を含むメゾスケールのゲノム構造の形成・機能原理をDNA物性的側面に着目して理解する。研究項目A03「ゲノムモダリティの制御と疾患」では、疾患・生理現象に関連する染色体レベルのマクロスケールのゲノム構造に対して、物理学に基づく形成・機能原理の理解を行う。